mRNA Processing
Eukaryotic pre-mRNA receives a 5′ cap and a 3′ poly (A) tail before introns are removed and the mRNA is considered ready for translation.
Pre-mRNA Processing
The eukaryotic pre-mRNA undergoes extensive processing before it is ready to be translated. The additional steps involved in eukaryotic mRNA maturation create a molecule with a much longer half-life than a prokaryotic mRNA. Eukaryotic mRNAs last for several hours, whereas the typical E. coli mRNA lasts no more than five seconds.
Pre-mRNAs are first coated in RNA-stabilizing proteins; these protect the pre-mRNA from degradation while it is processed and exported out of the nucleus. The three most important steps of pre-mRNA processing are the addition of stabilizing and signaling factors at the 5′ and 3′ ends of the molecule, and the removal of intervening sequences that do not specify the appropriate amino acids. In rare cases, the mRNA transcript can be “edited” after it is transcribed.
5′ Capping
While the pre-mRNA is still being synthesized, a 7-methylguanosine cap is added to the 5′ end of the growing transcript by a 5′-to-5′ phosphate linkage. This moiety protects the nascent mRNA from degradation. In addition, initiation factors involved in protein synthesis recognize the cap to help initiate translation by ribosomes.
5′ cap structure: Capping of the pre-mRNA involves the addition of 7-methylguanosine (m7G) to the 5′ end. The cap protects the 5′ end of the primary RNA transcript from attack by ribonucleases and is recognized by eukaryotic initiation factors involved in assembling the ribosome on the mature mRNA prior to initiating translation.
3′ Poly-A Tail
While RNA Polymerase II is still transcribing downstream of the proper end of a gene, the pre-mRNA is cleaved by an endonuclease-containing protein complex between an AAUAAA consensus sequence and a GU-rich sequence. This releases the functional pre-mRNA from the rest of the transcript, which is still attached to the RNA Polymerase. An enzyme called poly (A) polymerase (PAP) is part of the same protein complex that cleaves the pre-mRNA and it immediately adds a string of approximately 200 A nucleotides, called the poly (A) tail, to the 3′ end of the just-cleaved pre-mRNA. The poly (A) tail protects the mRNA from degradation, aids in the export of the mature mRNA to the cytoplasm, and is involved in binding proteins involved in initiating translation.
Poly (A) Polymerase adds a 3′ poly (A) tail to the pre-mRNA.: The pre-mRNA is cleaved off the rest of the growing transcript before RNA Polymerase II has stopped transcribing. This cleavage is done by an endonuclease-containing protein complex that binds to an AAUAAA sequence upstream of the cleavage site and to a GU-rich sequence downstream of the cut site. Immediately after the cleavage, Poly (A) Polymerase (PAP), which is also part of the protein complex, catalyzes the addition of up to 200 A nucleotides to the 3′ end of the just-cleaved pre-mRNA.
Pre-mRNA Splicing
Eukaryotic genes are composed of exons, which correspond to protein-coding sequences (ex-on signifies that they are expressed), and intervening sequences called introns (int-ron denotes their intervening role), which may be involved in gene regulation, but are removed from the pre-mRNA during processing. Intron sequences in mRNA do not encode functional proteins.
Discovery of Introns
The discovery of introns came as a surprise to researchers in the 1970s who expected that pre-mRNAs would specify protein sequences without further processing, as they had observed in prokaryotes. The genes of higher eukaryotes very often contain one or more introns. While these regions may correspond to regulatory sequences, the biological significance of having many introns or having very long introns in a gene is unclear. It is possible that introns slow down gene expression because it takes longer to transcribe pre-mRNAs with lots of introns. Alternatively, introns may be nonfunctional sequence remnants left over from the fusion of ancient genes throughout evolution. This is supported by the fact that separate exons often encode separate protein subunits or domains. For the most part, the sequences of introns can be mutated without ultimately affecting the protein product.
Intron Processing
All introns in a pre-mRNA must be completely and precisely removed before protein synthesis. If the process errs by even a single nucleotide, the reading frame of the rejoined exons would shift, and the resulting protein would be dysfunctional. The process of removing introns and reconnecting exons is called splicing. Introns are removed and degraded while the pre-mRNA is still in the nucleus. Splicing occurs by a sequence-specific mechanism that ensures introns will be removed and exons rejoined with the accuracy and precision of a single nucleotide. The splicing of pre-mRNAs is conducted by complexes of proteins and RNA molecules called spliceosomes.
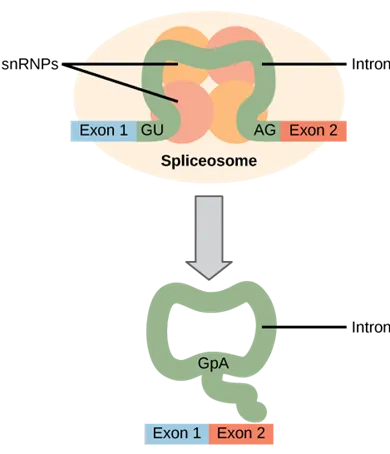
Pre-mRNA splicing: Pre-mRNA splicing involves the precise removal of introns from the primary RNA transcript. The splicing process is catalyzed by large complexes called spliceosomes. Each spliceosome is composed of five subunits called snRNPs. The spliceseome’s actions result in the splicing together of the two exons and the release of the intron in a lariat form.
Each spliceosome is composed of five subunits called snRNPs (for small nuclear ribonucleoparticles, and pronounced “snurps”.) Each snRNP is itself a complex of proteins and a special type of RNA found only in the nucleus called snRNAs (small nuclear RNAs). Spliceosomes recognize sequences at the 5′ end of the intron because introns always start with the nucleotides GU and they recognize sequences at the 3′ end of the intron because they always end with the nucleotides AG. The spliceosome cleaves the pre-mRNA’s sugar phosphate backbone at the G that starts the intron and then covalently attaches that G to an internal A nucleotide within the intron. Then the spliceosme connects the 3′ end of the first exon to the 5′ end of the following exon, cleaving the 3′ end of the intron in the process. This results in the splicing together of the two exons and the release of the intron in a lariat form.
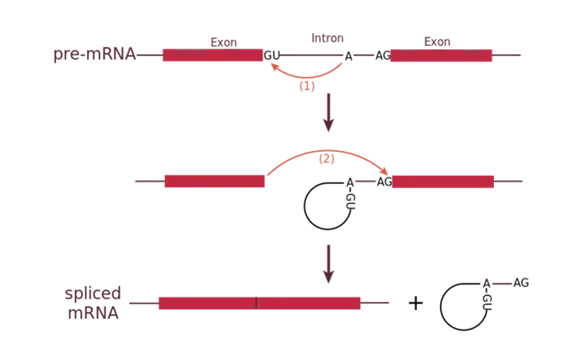
Mechanism of pre-mRNA splicing.: The snRNPs of the spliceosome were left out of this figure, but it shows the sites within the intron whose interactions are catalyzed by the spliceosome. Initially, the conserved G which starts an intron is cleaved from the 3′ end of the exon upstream to it and the G is covalently attached to an internal A within the intron. Then the 3′ end of the just-released exon is joined to the 5′ end of the next exon, cleaving the bond that attaches the 3′ end of the intron to its adjacent exon. This both joins the two exons and removes the intron in lariat form.
Processing of tRNAs and rRNAs
rRNA and tRNA are structural molecules that aid in protein synthesis but are not themselves translated into protein.
Processing of tRNAs and rRNAs
The tRNAs and rRNAs are structural molecules that have roles in protein synthesis; however, these RNAs are not themselves translated. In eukaryotes, pre-rRNAs are transcribed, processed, and assembled into ribosomes in the nucleolus, while pre-tRNAs are transcribed and processed in the nucleus and then released into the cytoplasm where they are linked to free amino acids for protein synthesis.
Ribosomal RNA (rRNA)
The four rRNAs in eukaryotes are first transcribed as two long precursor molecules. One contains just the pre-rRNA that will be processed into the 5S rRNA; the other spans the 28S, 5.8S, and 18S rRNAs. Enzymes then cleave the precursors into subunits corresponding to each rRNA. In bacteria, there are only three rRNAs and all are transcribed in one long precursor molecule that is cleaved into the individual rRNAs. Some of the bases of pre-rRNAs are methylated for added stability. Mature rRNAs make up 50-60% of each ribosome. Some of a ribosome’s RNA molecules are purely structural, whereas others have catalytic or binding activities.
The eukaryotic ribosome is composed of two subunits: a large subunit (60S) and a small subunit (40S). The 60S subunit is composed of the 28S rRNA, 5.8S rRNA, 5S rRNA, and 50 proteins. The 40S subunit is composed of the 18S rRNA and 33 proteins. The bacterial ribosome is composed of two similar subunits, with slightly different components. The bacterial large subunit is called the 50S subunit and is composed of the 23S rRNA, 5S rRNA, and 31 proteins, while the bacterial small subunit is called the 30S subunit and is composed of the 16S rRNA and 21 proteins.
The two subunits join to constitute a functioning ribosome that is capable of creating proteins.
Transfer RNA (tRNA)
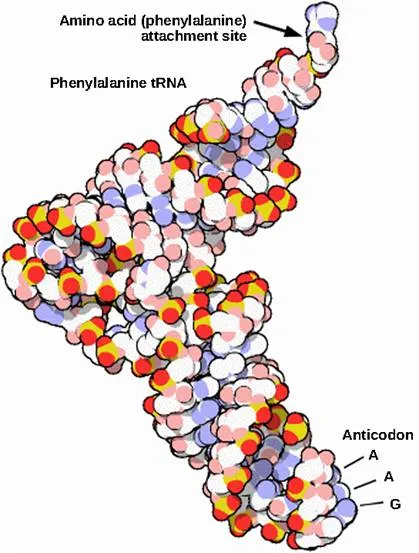
Each different tRNA binds to a specific amino acid and transfers it to the ribosome. Mature tRNAs take on a three-dimensional structure through intramolecular basepairing to position the amino acid binding site at one end and the anticodon in an unbasepaired loop of nucleotides at the other end. The anticodon is a three-nucleotide sequence, unique to each different tRNA, that interacts with a messenger RNA (mRNA) codon through complementary base pairing.
There are different tRNAs for the 21 different amino acids. Most amino acids can be carried by more than one tRNA.
Structure of tRNA: This is a space-filling model of a tRNA molecule that adds the amino acid phenylalanine to a growing polypeptide chain. The anticodon AAG binds the codon UUC on the mRNA. The amino acid phenylalanine is attached to the other end of the tRNA.