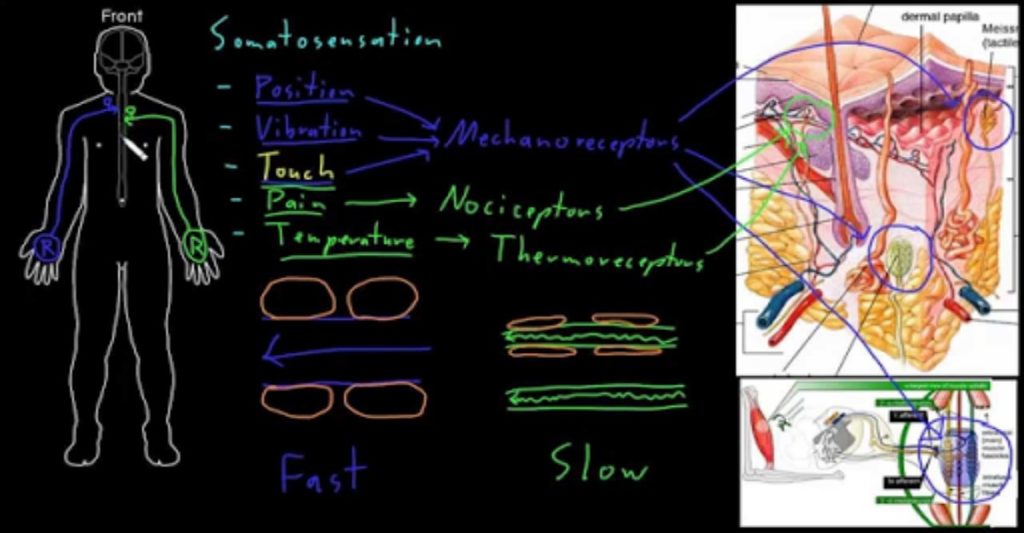
Somatosensation is a mixed sensory category and includes all sensation received from the skin and mucous membranes, as well from as the limbs and joints. Somatosensation is also known as tactile sense, or more familiarly, as the sense of touch. Somatosensation occurs all over the exterior of the body and at some interior locations as well. A variety of receptor types—embedded in the skin, mucous membranes, muscles, joints, internal organs, and cardiovascular system—play a role.
Recall that the epidermis is the outermost layer of skin in mammals. It is relatively thin, is composed of keratin-filled cells, and has no blood supply. The epidermis serves as a barrier to water and to invasion by pathogens. Below this, the much thicker dermis contains blood vessels, sweat glands, hair follicles, lymph vessels, and lipid-secreting sebaceous glands (Figure 17.4). Below the epidermis and dermis is the subcutaneous tissue, or hypodermis, the fatty layer that contains blood vessels, connective tissue, and the axons of sensory neurons. The hypodermis, which holds about 50 percent of the body’s fat, attaches the dermis to the bone and muscle, and supplies nerves and blood vessels to the dermis.
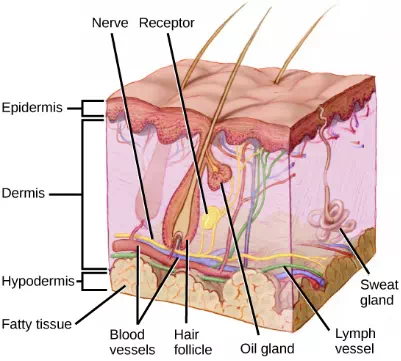
Figure 17.4. Mammalian skin has three layers: an epidermis, a dermis, and a hypodermis. (credit: modification of work by Don Bliss, National Cancer Institute)
Somatosensory Receptors
Sensory receptors are classified into five categories: mechanoreceptors, thermoreceptors, proprioceptors, pain receptors, and chemoreceptors. These categories are based on the nature of stimuli each receptor class transduces. What is commonly referred to as “touch” involves more than one kind of stimulus and more than one kind of receptor. Mechanoreceptors in the skin are described as encapsulated (that is, surrounded by a capsule) or unencapsulated (a group that includes free nerve endings). A free nerve ending, as its name implies, is an unencapsulated dendrite of a sensory neuron. Free nerve endings are the most common nerve endings in skin, and they extend into the middle of the epidermis. Free nerve endings are sensitive to painful stimuli, to hot and cold, and to light touch. They are slow to adjust to a stimulus and so are less sensitive to abrupt changes in stimulation.
There are three classes of mechanoreceptors: tactile, proprioceptors, and baroreceptors. Mechanoreceptors sense stimuli due to physical deformation of their plasma membranes. They contain mechanically gated ion channels whose gates open or close in response to pressure, touch, stretching, and sound.” There are four primary tactile mechanoreceptors in human skin: Merkel’s disks, Meissner’s corpuscles, Ruffini endings, and Pacinian corpuscle; two are located toward the surface of the skin and two are located deeper. A fifth type of mechanoreceptor, Krause end bulbs, are found only in specialized regions. Merkel’s disks (shown in Figure 17.5) are found in the upper layers of skin near the base of the epidermis, both in skin that has hair and on glabrous skin, that is, the hairless skin found on the palms and fingers, the soles of the feet, and the lips of humans and other primates. Merkel’s disks are densely distributed in the fingertips and lips. They are slow-adapting, unencapsulated nerve endings, and they respond to light touch. Light touch, also known as discriminative touch, is a light pressure that allows the location of a stimulus to be pinpointed. The receptive fields of Merkel’s disks are small with well-defined borders. That makes them finely sensitive to edges and they come into use in tasks such as typing on a keyboard.
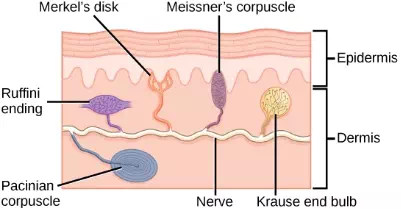
Figure 17.5. Four of the primary mechanoreceptors in human skin are shown. Merkel’s disks, which are unencapsulated, respond to light touch. Meissner’s corpuscles, Ruffini endings, Pacinian corpuscles, and Krause end bulbs are all encapsulated. Meissner’s corpuscles respond to touch and low-frequency vibration. Ruffini endings detect stretch, deformation within joints, and warmth. Pacinian corpuscles detect transient pressure and high-frequency vibration. Krause end bulbs detect cold.
Which of the following statements about mechanoreceptors is false?
1. Pacini corpuscles are found in both glabrous and hairy skin.
2. Merkel’s disks are abundant on the fingertips and lips.
3. Ruffini endings are encapsulated mechanoreceptors.
4. Meissner’s corpuscles extend into the lower dermis.
Meissner’s corpuscles, (shown in Figure 17.6) also known as tactile corpuscles, are found in the upper dermis, but they project into the epidermis. They, too, are found primarily in the glabrous skin on the fingertips and eyelids. They respond to fine touch and pressure, but they also respond to low-frequency vibration or flutter. They are rapidly adapting, fluid-filled, encapsulated neurons with small, well-defined borders and are responsive to fine details. Like Merkel’s disks, Meissner’s corpuscles are not as plentiful in the palms as they are in the fingertips.
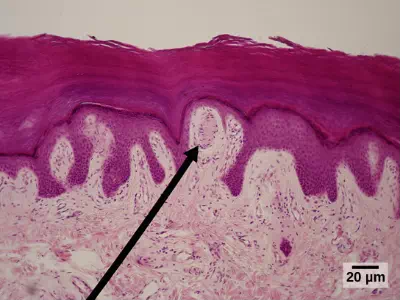
Figure 17.6. Meissner corpuscles in the fingertips, such as the one viewed here using bright field light microscopy, allow for touch discrimination of fine detail. (credit: modification of work by “Wbensmith”/Wikimedia Commons; scale-bar data from Matt Russell)
Deeper in the epidermis, near the base, are Ruffini endings, which are also known as bulbous corpuscles. They are found in both glabrous and hairy skin. These are slow-adapting, encapsulated mechanoreceptors that detect skin stretch and deformations within joints, so they provide valuable feedback for gripping objects and controlling finger position and movement. Thus, they also contribute to proprioception and kinesthesia. Ruffini endings also detect warmth. Note that these warmth detectors are situated deeper in the skin than are the cold detectors. It is not surprising, then, that humans detect cold stimuli before they detect warm stimuli.
Pacinian corpuscles (seen in Figure 17.7) are located deep in the dermis of both glabrous and hairy skin and are structurally similar to Meissner’s corpuscles; they are found in the bone periosteum, joint capsules, pancreas and other viscera, breast, and genitals. They are rapidly adapting mechanoreceptors that sense deep transient (but not prolonged) pressure and high-frequency vibration. Pacinian receptors detect pressure and vibration by being compressed, stimulating their internal dendrites. There are fewer Pacinian corpuscles and Ruffini endings in skin than there are Merkel’s disks and Meissner’s corpuscles.
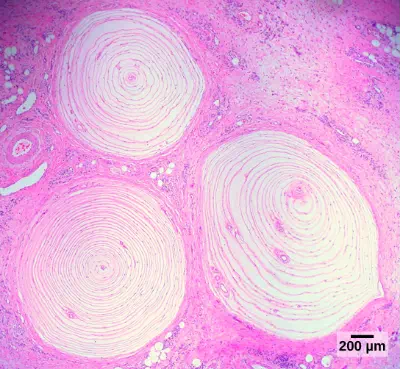
Figure 17.7. Pacinian corpuscles, such as these visualized using bright field light microscopy, detect pressure (touch) and high-frequency vibration. (credit: modification of work by Ed Uthman; scale-bar data from Matt Russell)
In proprioception, proprioceptive and kinesthetic signals travel through myelinated afferent neurons running from the spinal cord to the medulla. Neurons are not physically connected, but communicate via neurotransmitters secreted into synapses or “gaps” between communicating neurons. Once in the medulla, the neurons continue carrying the signals to the thalamus.
Muscle spindles are stretch receptors that detect the amount of stretch, or lengthening of muscles. Related to these are Golgi tendon organs, which are tension receptors that detect the force of muscle contraction. Proprioceptive and kinesthetic signals come from limbs. Unconscious proprioceptive signals run from the spinal cord to the cerebellum, the brain region that coordinates muscle contraction, rather than to the thalamus, like most other sensory information.
Barorecptors detect pressure changes in an organ. They are found in the walls of the carotid artery and the aorta where they monitor blood pressure, and in the lungs where they detect the degree of lung expansion. Stretch receptors are found at various sites in the digestive and urinary systems.
In addition to these two types of deeper receptors, there are also rapidly adapting hair receptors, which are found on nerve endings that wrap around the base of hair follicles. There are a few types of hair receptors that detect slow and rapid hair movement, and they differ in their sensitivity to movement. Some hair receptors also detect skin deflection, and certain rapidly adapting hair receptors allow detection of stimuli that have not yet touched the skin.
Integration of Signals from Mechanoreceptors
The configuration of the different types of receptors working in concert in human skin results in a very refined sense of touch. The nociceptive receptors—those that detect pain—are located near the surface. Small, finely calibrated mechanoreceptors—Merkel’s disks and Meissner’s corpuscles—are located in the upper layers and can precisely localize even gentle touch. The large mechanoreceptors—Pacinian corpuscles and Ruffini endings—are located in the lower layers and respond to deeper touch. (Consider that the deep pressure that reaches those deeper receptors would not need to be finely localized.) Both the upper and lower layers of the skin hold rapidly and slowly adapting receptors. Both primary somatosensory cortex and secondary cortical areas are responsible for processing the complex picture of stimuli transmitted from the interplay of mechanoreceptors.
Density of Mechanoreceptors
The distribution of touch receptors in human skin is not consistent over the body. In humans, touch receptors are less dense in skin covered with any type of hair, such as the arms, legs, torso, and face. Touch receptors are denser in glabrous skin (the type found on human fingertips and lips, for example), which is typically more sensitive and is thicker than hairy skin (4 to 5 mm versus 2 to 3 mm).
How is receptor density estimated in a human subject? The relative density of pressure receptors in different locations on the body can be demonstrated experimentally using a two-point discrimination test. In this demonstration, two sharp points, such as two thumbtacks, are brought into contact with the subject’s skin (though not hard enough to cause pain or break the skin). The subject reports if he or she feels one point or two points. If the two points are felt as one point, it can be inferred that the two points are both in the receptive field of a single sensory receptor. If two points are felt as two separate points, each is in the receptive field of two separate sensory receptors. The points could then be moved closer and re-tested until the subject reports feeling only one point, and the size of the receptive field of a single receptor could be estimated from that distance.
Thermoreception
In addition to Krause end bulbs that detect cold and Ruffini endings that detect warmth, there are different types of cold receptors on some free nerve endings: thermoreceptors, located in the dermis, skeletal muscles, liver, and hypothalamus, that are activated by different temperatures. Their pathways into the brain run from the spinal cord through the thalamus to the primary somatosensory cortex. Warmth and cold information from the face travels through one of the cranial nerves to the brain. You know from experience that a tolerably cold or hot stimulus can quickly progress to a much more intense stimulus that is no longer tolerable. Any stimulus that is too intense can be perceived as pain because temperature sensations are conducted along the same pathways that carry pain sensations
Pain
Pain is the name given to nociception, which is the neural processing of injurious stimuli in response to tissue damage. Pain is caused by true sources of injury, such as contact with a heat source that causes a thermal burn or contact with a corrosive chemical. But pain also can be caused by harmless stimuli that mimic the action of damaging stimuli, such as contact with capsaicins, the compounds that cause peppers to taste hot and which are used in self-defense pepper sprays and certain topical medications. Peppers taste “hot” because the protein receptors that bind capsaicin open the same calcium channels that are activated by warm receptors.
Nociception starts at the sensory receptors, but pain, inasmuch as it is the perception of nociception, does not start until it is communicated to the brain. There are several nociceptive pathways to and through the brain. Most axons carrying nociceptive information into the brain from the spinal cord project to the thalamus (as do other sensory neurons) and the neural signal undergoes final processing in the primary somatosensory cortex. Interestingly, one nociceptive pathway projects not to the thalamus but directly to the hypothalamus in the forebrain, which modulates the cardiovascular and neuroendocrine functions of the autonomic nervous system. Recall that threatening—or painful—stimuli stimulate the sympathetic branch of the visceral sensory system, readying a fight-or-flight response.
Summary
Somatosensation includes all sensation received from the skin and mucous membranes, as well as from the limbs and joints. Somatosensation occurs all over the exterior of the body and at some interior locations as well, and a variety of receptor types, embedded in the skin and mucous membranes, play a role.
There are several types of specialized sensory receptors. Rapidly adapting free nerve endings detect nociception, hot and cold, and light touch. Slowly adapting, encapsulated Merkel’s disks are found in fingertips and lips, and respond to light touch. Meissner’s corpuscles, found in glabrous skin, are rapidly adapting, encapsulated receptors that detect touch, low-frequency vibration, and flutter. Ruffini endings are slowly adapting, encapsulated receptors that detect skin stretch, joint activity, and warmth. Hair receptors are rapidly adapting nerve endings wrapped around the base of hair follicles that detect hair movement and skin deflection. Finally, Pacinian corpuscles are encapsulated, rapidly adapting receptors that detect transient pressure and high-frequency vibration.
Taste and Smell
Taste, also called gustation, and smell, also called olfaction, are the most interconnected senses in that both involve molecules of the stimulus entering the body and bonding to receptors. Smell lets an animal sense the presence of food or other animals—whether potential mates, predators, or prey—or other chemicals in the environment that can impact their survival. Similarly, the sense of taste allows animals to discriminate between types of foods. While the value of a sense of smell is obvious, what is the value of a sense of taste? Different tasting foods have different attributes, both helpful and harmful. For example, sweet-tasting substances tend to be highly caloric, which could be necessary for survival in lean times. Bitterness is associated with toxicity, and sourness is associated with spoiled food. Salty foods are valuable in maintaining homeostasis by helping the body retain water and by providing ions necessary for cells to function.
Tastes and Odors
Both taste and odor stimuli are molecules taken in from the environment. The primary tastes detected by humans are sweet, sour, bitter, salty and umami. The first four tastes need little explanation. The identification of umami as a fundamental taste occurred fairly recently—it was identified in 1908 by Japanese scientist Kikunae Ikeda while he worked with seaweed broth, but it was not widely accepted as a taste that could be physiologically distinguished until many years later. The taste of umami, also known as savoriness, is attributable to the taste of the amino acid L-glutamate. In fact, monosodium glutamate, or MSG, is often used in cooking to enhance the savory taste of certain foods. What is the adaptive value of being able to distinguish umami? Savory substances tend to be high in protein.
All odors that we perceive are molecules in the air we breathe. If a substance does not release molecules into the air from its surface, it has no smell. And if a human or other animal does not have a receptor that recognizes a specific molecule, then that molecule has no smell. Humans have about 350 olfactory receptor subtypes that work in various combinations to allow us to sense about 10,000 different odors. Compare that to mice, for example, which have about 1,300 olfactory receptor types, and therefore probably sense more odors. Both odors and tastes involve molecules that stimulate specific chemoreceptors. Although humans commonly distinguish taste as one sense and smell as another, they work together to create the perception of flavor. A person’s perception of flavor is reduced if he or she has congested nasal passages.