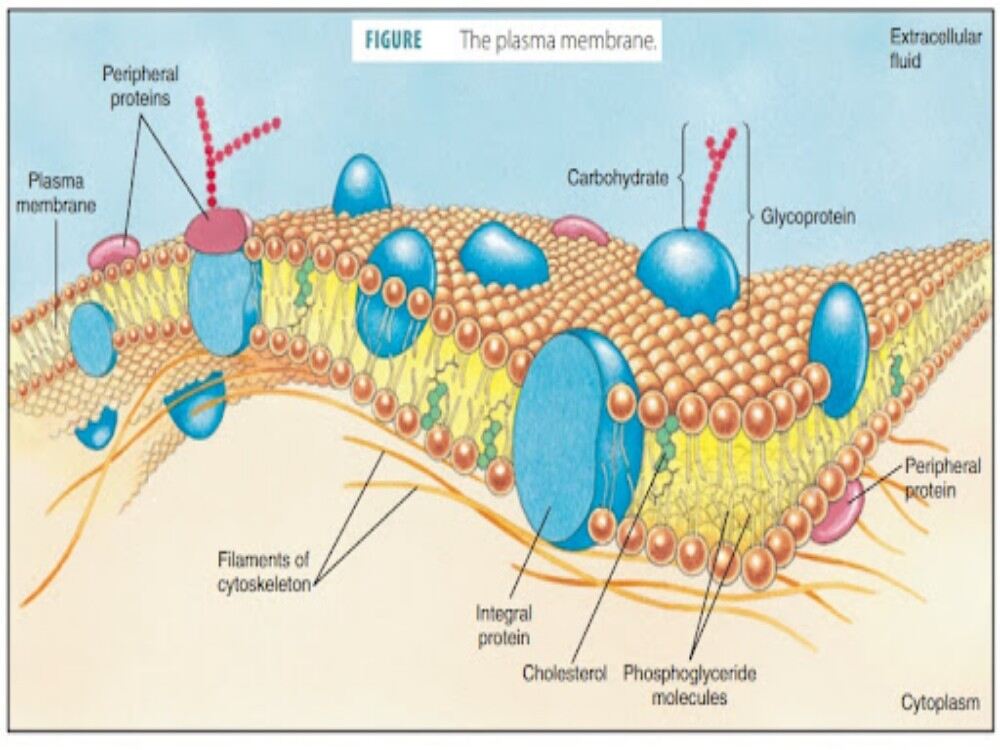
A cell’s plasma membrane defines the boundary of the cell and determines the nature of its contact with the environment. Cells exclude some substances, take in others, and excrete still others, all in controlled quantities. Plasma membranes enclose the borders of cells, but rather than being a static bag, they are dynamic and constantly in flux. The plasma membrane must be sufficiently flexible to allow certain cells, such as red blood cells and white blood cells, to change shape as they pass through narrow capillaries. These are the more obvious functions of a plasma membrane. In addition, the surface of the plasma membrane carries markers that allow cells to recognize one another, which is vital as tissues and organs form during early development, and which later plays a role in the “self” versus “non-self” distinction of the immune response.
The plasma membrane also carries receptors, which are attachment sites for specific substances that interact with the cell. Each receptor is structured to bind with a specific substance. For example, surface receptors of the membrane create changes in the interior, such as changes in enzymes of metabolic pathways. These metabolic pathways might be vital for providing the cell with energy, making specific substances for the cell, or breaking down cellular waste or toxins for disposal. Receptors on the plasma membrane’s exterior surface interact with hormones or neurotransmitters, and allow their messages to be transmitted into the cell. Some recognition sites are used by viruses as attachment points. Although they are highly specific, pathogens like viruses may evolve to exploit receptors to gain entry to a cell by mimicking the specific substance that the receptor is meant to bind. This specificity helps to explain why human immunodeficiency virus (HIV) or any of the five types of hepatitis viruses invade only specific cells.
Fluid Mosaic Model
In 1972, S. J. Singer and Garth L. Nicolson proposed a new model of the plasma membrane that, compared to earlier understanding, better explained both microscopic observations and the function of the plasma membrane. This was called the fluid mosaic model. The model has evolved somewhat over time, but still best accounts for the structure and functions of the plasma membrane as we now understand them. The fluid mosaic model describes the structure of the plasma membrane as a mosaic of components—including phospholipids, cholesterol, proteins, and carbohydrates—in which the components are able to flow and change position, while maintaining the basic integrity of the membrane. Both phospholipid molecules and embedded proteins are able to diffuse rapidly and laterally in the membrane. The fluidity of the plasma membrane is necessary for the activities of certain enzymes and transport molecules within the membrane. Plasma membranes range from 5–10 nm thick. As a comparison, human red blood cells, visible via light microscopy, are approximately 8 µm thick, or approximately 1,000 times thicker than a plasma membrane.
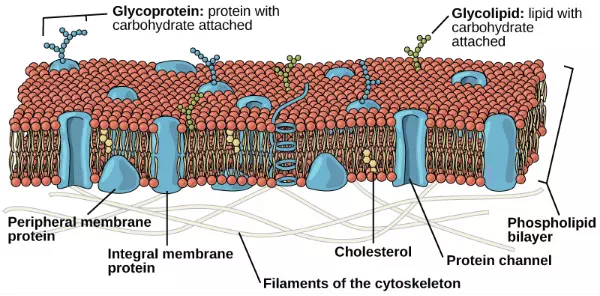
The fluid mosaic model of the plasma membrane structure describes the plasma membrane as a fluid combination of phospholipids, cholesterol, proteins, and carbohydrates.
The plasma membrane is made up primarily of a bilayer of phospholipids with embedded proteins, carbohydrates, glycolipids, and glycoproteins, and, in animal cells, cholesterol. The amount of cholesterol in animal plasma membranes regulates the fluidity of the membrane and changes based on the temperature of the cell’s environment. In other words, cholesterol acts as antifreeze in the cell membrane and is more abundant in animals that live in cold climates.
The main fabric of the membrane is composed of two layers of phospholipid molecules, and the polar ends of these molecules (which look like a collection of balls in an artist’s rendition of the model) are in contact with aqueous fluid both inside and outside the cell. Thus, both surfaces of the plasma membrane are hydrophilic. In contrast, the interior of the membrane, between its two surfaces, is a hydrophobic or nonpolar region because of the fatty acid tails. This region has no attraction for water or other polar molecules.
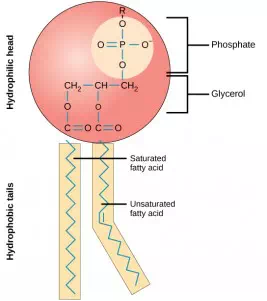
This phospholipid molecule is composed of a hydrophilic head and two hydrophobic tails. The hydrophilic head group consists of a phosphate-containing group attached to a glycerol molecule. The hydrophobic tails, each containing either a saturated or an unsaturated fatty acid, are long hydrocarbon chains.
Proteins make up the second major chemical component of plasma membranes. Integral proteins are embedded in the plasma membrane and may span all or part of the membrane. Integral proteins may serve as channels or pumps to move materials into or out of the cell. Peripheral proteins are found on the exterior or interior surfaces of membranes, attached either to integral proteins or to phospholipid molecules. Both integral and peripheral proteins may serve as enzymes, as structural attachments for the fibers of the cytoskeleton, or as part of the cell’s recognition sites.
Carbohydrates are the third major component of plasma membranes. They are always found on the exterior surface of cells and are bound either to proteins (forming glycoproteins) or to lipids (forming glycolipids). These carbohydrate chains may consist of 2–60 monosaccharide units and may be either straight or branched. Along with peripheral proteins, carbohydrates form specialized sites on the cell surface that allow cells to recognize each other.
Evolution in Action
How Viruses Infect Specific OrgansSpecific glycoprotein molecules exposed on the surface of the cell membranes of host cells are exploited by many viruses to infect specific organs. For example, HIV is able to penetrate the plasma membranes of specific kinds of white blood cells called T-helper cells and monocytes, as well as some cells of the central nervous system. The hepatitis virus attacks only liver cells.
These viruses are able to invade these cells, because the cells have binding sites on their surfaces that the viruses have exploited with equally specific glycoproteins in their coats. (Figure 3.23). The cell is tricked by the mimicry of the virus coat molecules, and the virus is able to enter the cell. Other recognition sites on the virus’s surface interact with the human immune system, prompting the body to produce antibodies. Antibodies are made in response to the antigens (or proteins associated with invasive pathogens). These same sites serve as places for antibodies to attach, and either destroy or inhibit the activity of the virus. Unfortunately, these sites on HIV are encoded by genes that change quickly, making the production of an effective vaccine against the virus very difficult. The virus population within an infected individual quickly evolves through mutation into different populations, or variants, distinguished by differences in these recognition sites. This rapid change of viral surface markers decreases the effectiveness of the person’s immune system in attacking the virus, because the antibodies will not recognize the new variations of the surface patterns.
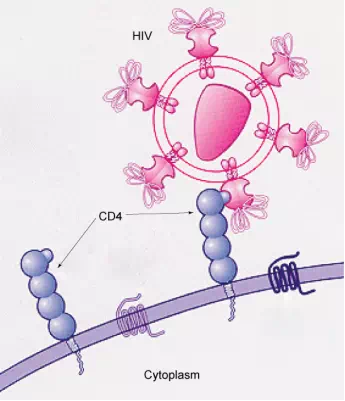
Figure 3.23 HIV docks at and binds to the CD4 receptor, a glycoprotein on the surface of T cells, before entering, or infecting, the cell. (credit: modification of work by US National Institutes of Health/National Institute of Allergy and Infectious Diseases.
Summary
The modern understanding of the plasma membrane is referred to as the fluid mosaic model. The plasma membrane is composed of a bilayer of phospholipids, with their hydrophobic, fatty acid tails in contact with each other. The landscape of the membrane is studded with proteins, some of which span the membrane. Some of these proteins serve to transport materials into or out of the cell. Carbohydrates are attached to some of the proteins and lipids on the outward-facing surface of the membrane. These form complexes that function to identify the cell to other cells. The fluid nature of the membrane owes itself to the configuration of the fatty acid tails, the presence of cholesterol embedded in the membrane (in animal cells), and the mosaic nature of the proteins and protein-carbohydrate complexes, which are not firmly fixed in place. Plasma membranes enclose the borders of cells, but rather than being a static bag, they are dynamic and constantly in flux.
Passive Transport
Plasma membranes must allow certain substances to enter and leave a cell, while preventing harmful material from entering and essential material from leaving. In other words, plasma membranes are selectively permeable—they allow some substances through but not others. If they were to lose this selectivity, the cell would no longer be able to sustain itself, and it would be destroyed. Some cells require larger amounts of specific substances than do other cells; they must have a way of obtaining these materials from the extracellular fluids. This may happen passively, as certain materials move back and forth, or the cell may have special mechanisms that ensure transport. Most cells expend most of their energy, in the form of adenosine triphosphate (ATP), to create and maintain an uneven distribution of ions on the opposite sides of their membranes. The structure of the plasma membrane contributes to these functions, but it also presents some problems.
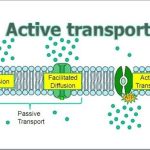
The most direct forms of membrane transport are passive. Passive transport is a naturally occurring phenomenon and does not require the cell to expend energy to accomplish the movement. In passive transport, substances move from an area of higher concentration to an area of lower concentration in a process called diffusion. A physical space in which there is a different concentration of a single substance is said to have a concentration gradient.
Selective Permeability
Plasma membranes are asymmetric, meaning that despite the mirror image formed by the phospholipids, the interior of the membrane is not identical to the exterior of the membrane. Integral proteins that act as channels or pumps work in one direction. Carbohydrates, attached to lipids or proteins, are also found on the exterior surface of the plasma membrane. These carbohydrate complexes help the cell bind substances that the cell needs in the extracellular fluid. This adds considerably to the selective nature of plasma membranes.
Recall that plasma membranes have hydrophilic and hydrophobic regions. This characteristic helps the movement of certain materials through the membrane and hinders the movement of others. Lipid-soluble material can easily slip through the hydrophobic lipid core of the membrane. Substances such as the fat-soluble vitamins A, D, E, and K readily pass through the plasma membranes in the digestive tract and other tissues. Fat-soluble drugs also gain easy entry into cells and are readily transported into the body’s tissues and organs. Molecules of oxygen and carbon dioxide have no charge and pass through by simple diffusion.
Polar substances, with the exception of water, present problems for the membrane. While some polar molecules connect easily with the outside of a cell, they cannot readily pass through the lipid core of the plasma membrane. Additionally, whereas small ions could easily slip through the spaces in the mosaic of the membrane, their charge prevents them from doing so. Ions such as sodium, potassium, calcium, and chloride must have a special means of penetrating plasma membranes. Simple sugars and amino acids also need help with transport across plasma membranes.
Diffusion
Diffusion is a passive process of transport. A single substance tends to move from an area of high concentration to an area of low concentration until the concentration is equal across the space. You are familiar with diffusion of substances through the air. For example, think about someone opening a bottle of perfume in a room filled with people. The perfume is at its highest concentration in the bottle and is at its lowest at the edges of the room. The perfume vapor will diffuse, or spread away, from the bottle, and gradually, more and more people will smell the perfume as it spreads. Materials move within the cell’s cytosol by diffusion, and certain materials move through the plasma membrane by diffusion (Figure 3.24). Diffusion expends no energy. Rather the different concentrations of materials in different areas are a form of potential energy, and diffusion is the dissipation of that potential energy as materials move down their concentration gradients, from high to low.
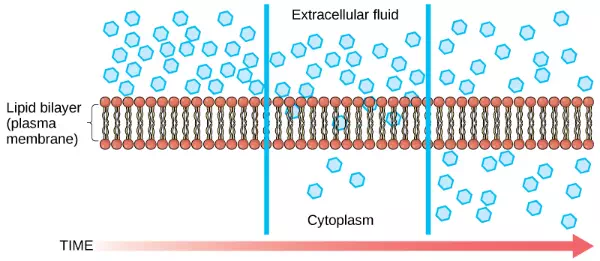
Figure 3.24 Diffusion through a permeable membrane follows the concentration gradient of a substance, moving the substance from an area of high concentration to one of low concentration. (credit: modification of work by Mariana Ruiz Villarreal)
Each separate substance in a medium, such as the extracellular fluid, has its own concentration gradient, independent of the concentration gradients of other materials. Additionally, each substance will diffuse according to that gradient.
Several factors affect the rate of diffusion.
· Extent of the concentration gradient: The greater the difference in concentration, the more rapid the diffusion. The closer the distribution of the material gets to equilibrium, the slower the rate of diffusion becomes.
· Mass of the molecules diffusing: More massive molecules move more slowly, because it is more difficult for them to move between the molecules of the substance they are moving through; therefore, they diffuse more slowly.
· Temperature: Higher temperatures increase the energy and therefore the movement of the molecules, increasing the rate of diffusion.
· Solvent density: As the density of the solvent increases, the rate of diffusion decreases. The molecules slow down because they have a more difficult time getting through the denser medium.
Facilitated transport
In facilitated transport, also called facilitated diffusion, material moves across the plasma membrane with the assistance of transmembrane proteins down a concentration gradient (from high to low concentration) without the expenditure of cellular energy. However, the substances that undergo facilitated transport would otherwise not diffuse easily or quickly across the plasma membrane. The solution to moving polar substances and other substances across the plasma membrane rests in the proteins that span its surface. The material being transported is first attached to protein or glycoprotein receptors on the exterior surface of the plasma membrane. This allows the material that is needed by the cell to be removed from the extracellular fluid. The substances are then passed to specific integral proteins that facilitate their passage, because they form channels or pores that allow certain substances to pass through the membrane. The integral proteins involved in facilitated transport are collectively referred to as transport proteins, and they function as either channels for the material or carriers.
Osmosis
Osmosis is the diffusion of water through a semipermeable membrane according to the concentration gradient of water across the membrane. Whereas diffusion transports material across membranes and within cells, osmosis transports only water across a membrane and the membrane limits the diffusion of solutes in the water. Osmosis is a special case of diffusion. Water, like other substances, moves from an area of higher concentration to one of lower concentration. Imagine a beaker with a semipermeable membrane, separating the two sides or halves . On both sides of the membrane, the water level is the same, but there are different concentrations on each side of a dissolved substance, or solute, that cannot cross the membrane. If the volume of the water is the same, but the concentrations of solute are different, then there are also different concentrations of water, the solvent, on either side of the membrane.
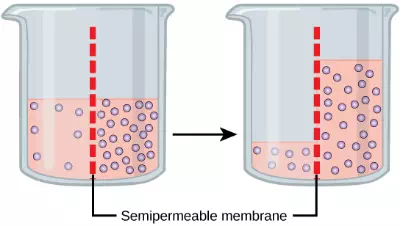
Figure 3.25 In osmosis, water always moves from an area of higher concentration (of water) to one of lower concentration (of water). In this system, the solute cannot pass through the selectively permeable membrane.
A principle of diffusion is that the molecules move around and will spread evenly throughout the medium if they can. However, only the material capable of getting through the membrane will diffuse through it. In this example, the solute cannot diffuse through the membrane, but the water can. Water has a concentration gradient in this system. Therefore, water will diffuse down its concentration gradient, crossing the membrane to the side where it is less concentrated. This diffusion of water through the membrane—osmosis—will continue until the concentration gradient of water goes to zero. Osmosis proceeds constantly in living systems.
Tonicity
Tonicity describes the amount of solute in a solution. The measure of the tonicity of a solution, or the total amount of solutes dissolved in a specific amount of solution, is called its osmolarity. Three terms—hypotonic, isotonic, and hypertonic—are used to relate the osmolarity of a cell to the osmolarity of the extracellular fluid that contains the cells. In a hypotonic solution, such as tap water, the extracellular fluid has a lower concentration of solutes than the fluid inside the cell, and water enters the cell. (In living systems, the point of reference is always the cytoplasm, so the prefix hypo– means that the extracellular fluid has a lower concentration of solutes, or a lower osmolarity, than the cell cytoplasm.) It also means that the extracellular fluid has a higher concentration of water than does the cell. In this situation, water will follow its concentration gradient and enter the cell. This may cause an animal cell to burst, or lyse.
In a hypertonic solution (the prefix hyper– refers to the extracellular fluid having a higher concentration of solutes than the cell’s cytoplasm), the fluid contains less water than the cell does, such as seawater. Because the cell has a lower concentration of solutes, the water will leave the cell. In effect, the solute is drawing the water out of the cell. This may cause an animal cell to shrivel, or crenate.
In an isotonic solution, the extracellular fluid has the same osmolarity as the cell. If the concentration of solutes of the cell matches that of the extracellular fluid, there will be no net movement of water into or out of the cell. Blood cells in hypertonic, isotonic, and hypotonic solutions take on characteristic appearances (Figure 3.26).
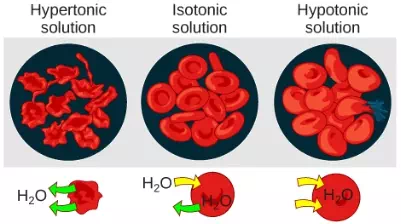
Figure 3.26 Osmotic pressure changes the shape of red blood cells in hypertonic, isotonic, and hypotonic solutions. (credit: modification of work by Mariana Ruiz Villarreal)
A doctor injects a patient with what the doctor thinks is isotonic saline solution. The patient dies, and autopsy reveals that many red blood cells have been destroyed. Do you think the solution the doctor injected was really isotonic?
<!– No, it must have been hypotonic, as a hypotonic solution would cause water to enter the cells, thereby making them burst. –>
Some organisms, such as plants, fungi, bacteria, and some protists, have cell walls that surround the plasma membrane and prevent cell lysis. The plasma membrane can only expand to the limit of the cell wall, so the cell will not lyse. In fact, the cytoplasm in plants is always slightly hypertonic compared to the cellular environment, and water will always enter a cell if water is available. This influx of water produces turgor pressure, which stiffens the cell walls of the plant . In nonwoody plants, turgor pressure supports the plant. If the plant cells become hypertonic, as occurs in drought or if a plant is not watered adequately, water will leave the cell. Plants lose turgor pressure in this condition and wilt.
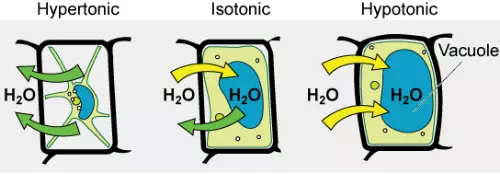
Figure 3.27 The turgor pressure within a plant cell depends on the tonicity of the solution that it is bathed in. (credit: modification of work by Mariana Ruiz Villarreal)
Comments are closed